Suppression of Epithelial-Mesenchymal Transition by Prostaglandin E2 in Retinal Pigment Epithelial Cells
Orita T, Kimura K*, Kohata Y, Sakuma A, Fujitsu Y, Wakuta M, Sonoda KH
Department of Ophthalmology, Yamaguchi University Graduate School of Medicine, 1-1-1 Minami-Kogushi, Ube City, Yamaguchi 755-8505, Japan.
*Corresponding Author
Kazuhiro Kimura,M.D., Ph.D.,
Department of Ophthalmology,
Yamaguchi University Graduate School of Medicine,
1-1-1 Minami-Kogushi, Ube City, Yamaguchi 755-8505, Japan.
Tel.: +81-836-22-2278
Fax: +81-836-22-2334
E-mail: k.kimura@yamaguchi-u.ac.jp
Received: March 18, 2016; Accepted: April 13, 2016; Published: April 19, 2016.
Citation: Kimura K et al., (2016) Suppression of Epithelial-Mesenchymal Transition by Prostaglandin E2 in Retinal Pigment Epithelial Cells. Int J Ophthalmol Eye Res. 4(3), 192-197.DOI : dx.doi.org/10.19070/2332-290X-1500039
Copyright: Kimura K© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background/Aims: An epithelial to mesenchymal transition of retinal pigment epithelium (RPE) cells plays an important role in the formation of proliferative fibrocellular membranes in individuals with proliferative retinopathy. Prostaglandin E2 (PGE2) exhibits various physiological actions including both pro- and anti-inflammatory effects. We have now investigated the effect of PGE2 on the epithelial-mesenchymal transition (EMT) of mouse RPE cells and fibrotic reactions in a type I collagen gel.
Methods: Mouse RPE cells were incubated in three dimentional type 1 collagen gel culture system with transforming growth factor (TGF)-β2 (1ng/ml) in the absence or presence of PGE2 at various concentration. The expression of α-smooth muscle actin as well as matrix metalloproteinase (MMP)-3, the phosphorylation of Smad2, paxillin were examined by immunoblot analysis. The release of MMP-2 and MMP-9 was examined by gelatin zymography.
Results: PGE2 inhibited the TGF-β2–induced contraction of collagen gels mediated by RPE cells in concentration dependent manner. The expression of α–smooth muscle actin and fibronectin as mesenchymal markers was inhibited. The release of MMP–2 and MMP-9 as well as the phosphorylation of Smad2 and paxillin induced by TGF-β2 in RPE cells were also attenuated by PGE2.
Conclusion: PGE2 may inhibit the contraction of proliferative fibrocellular membranes that can lead to retinal detachment in individuals with progressive vitreoretinal diseases.
2.Introduction
3.Material and Methods
3.1.Materials
3.2.Isolation and Culture of Mouse RPE Cells
3.3.Assay of Collagen Gel Contraction
3.4.Immunoblot Analysis
3.5.Gelatin Zymography
3.6.Statistical Analysis
4.Results
4.1.Effect of PGE2 on TGF-β2-Induced Collagen Gel Contraction Mediated by RPE Cells
4.2.Effect of PGE2 on Smad2 Phosphorylation Induced by TGF-β2 in RPE Cells
4.3.Effect of PGE2 on the Expression of α-SMA Induced by TGF-β2 in RPE Cells
4.3.Effect of PGE2 on Paxillin Phosphorylation Induced by TGF-β2 in RPE Cells
4.4.Effect of PGE2 on MMP release Induced by TGF-β2 in RPE Cells
5.Discussion
6.Acknowledgements
7.References
Keywords
Retinal Pigment Epithelial Cell; TGF-Β2; Epithelial-Mesenchymal Transition; Collagen Gel Contraction; Prostaglandin E2.
Introduction
Proliferative ocular diseases such as proliferative vitreoretinopathy (PVR) and proliferative diabetic retinopathy (PDR) can result in visual impairment or blindness. The contraction of proliferative tissue that has formed on or under the retina gives rise to retinal tearing and detachment and may be responsible for failure of surgery to repair retinal damage. The proliferative tissue consists of retinal pigment epithelium (RPE) cells, glial cells, and fibroblasts and is exposed to growth factors and inflammatory cytokines such as platelet-derived growth factor (PDGF), transforming growth factor–β (TGF-β), and hepatocyte growth factor (HGF). In particular, RPE cells that have undergone the epithelial-mesenchymal transition (EMT) play a key role in the formation of proliferative fibrocellular membranes [1]. These cells express α–smooth muscle actin (α-SMA), fibronectin, and vimentin, all of which are markers of the EMT, as well as matrix metalloproteinases (MMPs) and connective tissue growth factor (CTGF) [2]. Signaling by TGF-β and Smad proteins regulates the EMT, with the phosphorylation of Smad2 and Smad3 induced by TGF-β leading to the transcriptional activation of genes related to this phenotypic change. Mechanical forces generated by mesenchymal cells are required for the assembly of focal adhesion proteins such as focal adhesion kinase, paxillin, and vinculin.
Prostaglandin E2 (PGE2) is produced widely throughout the body and has various biological effects. In particular, it plays a central role in the regulation of inflammation. PGE2 exerts its effects through binding to E-prostanoid (EP) receptors, of which there are four subtypes (EP1, EP2, EP3, and EP4) [3]. These four subtypes of EP receptor are all members of the G protein–coupled receptor family, but they differ in the intracellular signaling pathways to which they are linked [4]. Most clinically beneficial effects of PGE2 are mediated through EP2 and EP4 receptors [5]. PGE2 mediates bronchodilation via the EP2 receptor and most of its anti-inflammatory effects are mediated through EP2 or EP4 [6]. EP2 and EP4 receptors have also been found to be expressed in proliferative tissue in the retina [7].
Inflammatory pathways activated by PGE2 have also been implicated in the formation of PVR membranes [8]. The EP4 receptor promotes the production of vascular endothelial growth factor (VEGF) and is thereby thought to play an important role in pathological ocular angiogenesis characteristic of neovascular retinopathies such as PDR and age-related macular degeneration [9].
PGE2 exerts both positive and negative effects on inflammation. It thus promotes inflammation induced by leukotrienes as well as inhibits the release of inflammatory mediators by monocytesmacrophages and dendritic cells [10]. We have now examined the effects of PGE2 on collagen gel contraction mediated by mouse RPE cells exposed to TGF-β in vitro as a model for the contraction of proliferative fibrocellular membranes in PVR and PDR.
Material and Methods
Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum, and trypsin-EDTA were obtained from Invitrogen-Gibco (Carlsbad, CA), and 24-well culture plates and 60-mm culture dishes were from Corning-Costar (Corning, NY). Native porcine type I collagen (acid solubilized) and reconstitution buffer were obtained from Nitta Gelatin (Osaka, Japan); bovine serum albumin (BSA) was from Nacalai Tesque (Kyoto, Japan); a protease inhibitor cocktail was from Sigma-Aldrich (St. Louis, MO). PGE2 was obtained from Biomol (Plymouth Meeting, PA), and recombinant human TGF-β2 was from R&D Systems (Minneapolis, MN). Mouse monoclonal antibodies to α-SMA were from Sigma-Aldrich, and those to Smad2, to phosphorylated Smad2, and to phosphorylated paxillin were from Cell Signaling (Beverly, MA). Mouse antibodies to paxillin was from BD Biosciences Pharmingen (San Diego, CA) and those to MMP- 3 was from Assay Biotechnology (Sunnyvale, CA). Horseradish peroxidase–conjugated goat secondary antibodies and ECL Plus detection reagents were from Amersham Biosciences (Little Chalfont, UK).
Mouse RPE cells were isolated as described previously [11]. The cells were maintained under a humidified atmosphere of 5% CO2 at 37°C in culture dishes containing DMEM supplemented with 10% fetal bovine serum.
Collagen gels were prepared as described previously [12]. In brief, 24-well culture plates were coated with 1% BSA for 1 h at 37°C. Mouse RPE cells were harvested by exposure to trypsin- EDTA, washed twice with serum-free DMEM, and resuspended in the same medium. Type I collagen (3 mg/ml), 10× DMEM, reconstitution buffer, mouse RPE cell suspension (1.1 × 107cells/ ml in DMEM), and deionized water were mixed on ice in a volume ratio of 7:1:1:0.2:1.8 (final concentration of type I collagen, 1.9 mg/ml; final cell density, 2 × 105/ml). A portion (0.5 ml) of the resulting mixture was added to each BSA-coated well of the culture plates and allowed to solidify by incubation at 37°C under 5% CO2 for 30 min. The collagen gels were freed from the sides of the wells with a microspatula, and serum-free DMEM (0.5 ml) with or without TGF-β2 (1 ng/ml) and PGE2 (30 or 100 μM) was then added on top of each gel. The cells were then cultured for 24 h, after which the diameter of each gel was measured with a ruler for determination of the extent of contraction.
Cells in collagen gels were lysed in a solution containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM EDTA, 5 mM NaF, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM Na3VO4, and 1% protease inhibitor cocktail, the lysates were subjected to SDS-polyacrylamide gel electrophoresis on a 10% gel, and the separated proteins were transferred electrophoretically to a nitrocellulose membrane. Nonspecific sites of the membrane were blocked, and it was then incubated consecutively with primary antibodies, horseradish peroxidase–conjugated secondary antibodies, and ECL reagents.
Gelatin zymography was performed as described previously [12]. In brief, culture supernatants (8 μl) from collagen gel incubations were mixed with 4 μl of nonreducing SDS sample buffer [125 mM Tris-HCl (pH 6.8), 20% glycerol, 2% SDS, 0.002% bromophenol blue], and 5 μl of the resulting mixture were subjected to SDSpolyacrylamide gel electrophoresis in the dark at 4°C on a 10% gel containing 0.1% gelatin. The gel was then washed with 2.5% Triton X-100 for 1 h before incubation for 18 h at 37°C in a reaction mixture containing 50 mM Tris-HCl (pH 7.5), 5 mM CaCl2, and 1% Triton X-100. The gel was finally stained with Coomassie brilliant blue.
Quantitative data are presented as means ± SD and were analyzed by Dunnett’s test. A P value of <0.05 was considered statistically significant.
Results
We first examined the possible effect of PGE2 on TGF-β2– induced collagen gel contraction mediated by RPE cells. The cells were thus incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 and PGE2 at various concentration for 24 h. The stimulatory effect of TGF-β2 on collagen gel contraction mediated by RPE cells was inhibited by PGE2 in a concentration-dependent manner (Figure 1).
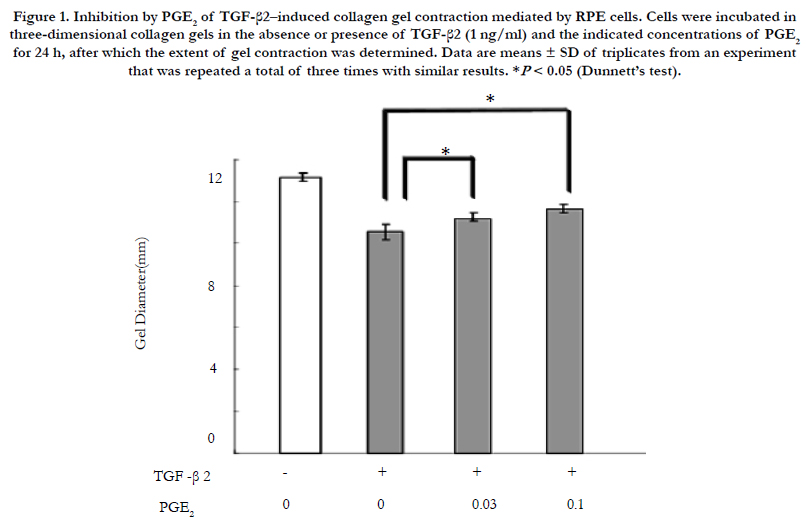
Figure 1. Inhibition by PGE2 of TGF-β2–induced collagen gel contraction mediated by RPE cells. Cells were incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 (1 ng/ml) and the indicated concentrations of PGE2 for 24 h, after which the extent of gel contraction was determined. Data are means ± SD of triplicates from an experiment that was repeated a total of three times with similar results. *P < 0.05 (Dunnett’s test).
To investigate the mechanism by which PGE2 inhibited TGF-β2– induced collagen gel contraction mediated by RPE cells, we examined the effect of this agent on the phosphorylation status of Smad2. Cells were incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 and PGE2 for 24 h, after which cell lysates were prepared and subjected to immunoblot analysis. The TGF-β2–induced phosphorylation of Smad2 was inhibited by PGE2 (Figure 2).
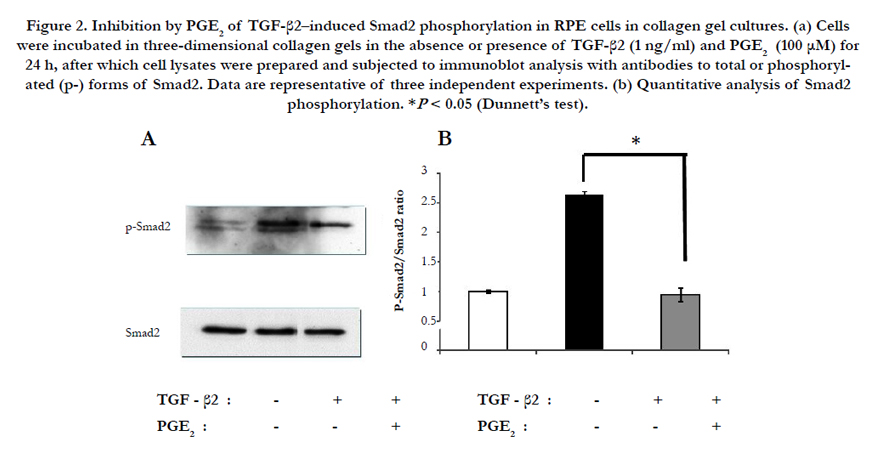
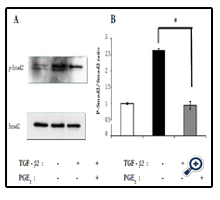
Figure 2. Inhibition by PGE2 of TGF-β2–induced Smad2 phosphorylation in RPE cells in collagen gel cultures. (a) Cells were incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 (1 ng/ml) and PGE2 (100 μM) for 24 h, after which cell lysates were prepared and subjected to immunoblot analysis with antibodies to total or phosphorylated (p-) forms of Smad2. Data are representative of three independent experiments. (b) Quantitative analysis of Smad2 phosphorylation. *P < 0.05 (Dunnett’s test).
We next examined the effect of PGE2 on expression of the mesenchymal marker α-SMA induced by TGF-β2 in RPE cells. Immunoblot analysis of cells incubated in collagen gels for 24 h in the absence or presence of TGF-β2 and PGE2 revealed that the up-regulation of α-SMA expression induced by TGF-β2 was inhibited by PGE2 (Figure 3).
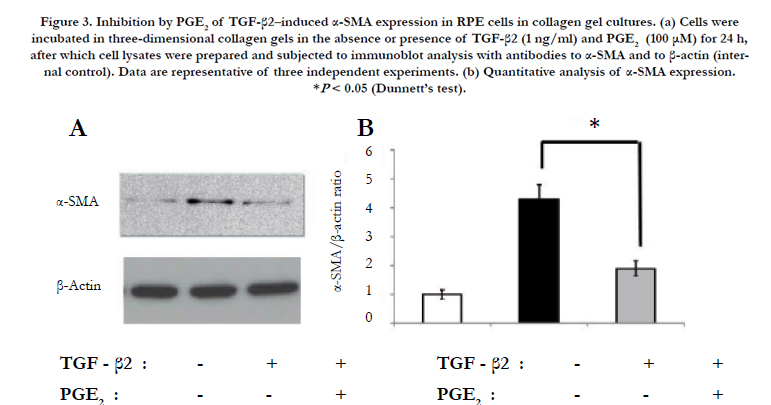
Figure 3. Inhibition by PGE2 of TGF-β2–induced α-SMA expression in RPE cells in collagen gel cultures. (a) Cells were incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 (1 ng/ml) and PGE2 (100 μM) for 24 h, after which cell lysates were prepared and subjected to immunoblot analysis with antibodies to α-SMA and to β-actin (internal control). Data are representative of three independent experiments. (b) Quantitative analysis of α-SMA expression. *P < 0.05 (Dunnett’s test).
The effect of PGE2 on phosphorylation of the focal adhesion protein paxillin induced by TGF-β2 was examined in RPE cells in collagen gel cultures. Incubation of the cells for 24 h with TGF-β2 increased the level of paxillin phosphorylation, and this effect was prevented by PGE2 (Figure 4).
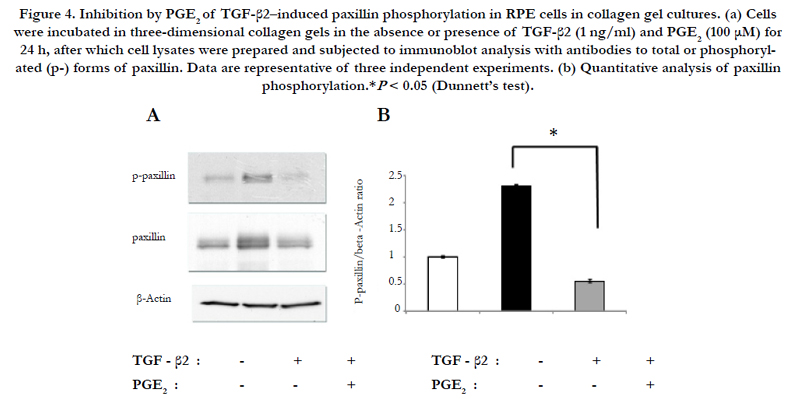
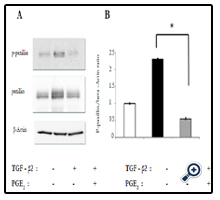
Figure 4. Inhibition by PGE2 of TGF-β2–induced paxillin phosphorylation in RPE cells in collagen gel cultures. (a) Cells were incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 (1 ng/ml) and PGE2 (100 μM) for 24 h, after which cell lysates were prepared and subjected to immunoblot analysis with antibodies to total or phosphorylated (p-) forms of paxillin. Data are representative of three independent experiments. (b) Quantitative analysis of paxillin phosphorylation.*P < 0.05 (Dunnett’s test).
Culture supernatants obtained from RPE cells incubated in collagen gels in the absence or presence of TGF-β2 and PGE2 for 24 h were analyzed for the release of MMPs. Gelatin zymography of the culture supernatants revealed that TGF-β2 increased the production of both MMP-2 and MMP-9 by RPE cells, and that these effects were inhibited by PGE2 (Figure 5). Immunoblot analysis revealed that neither TGF-β2 nor PGE2 affected the release of MMP-3 by RPE cells (Figure 5).
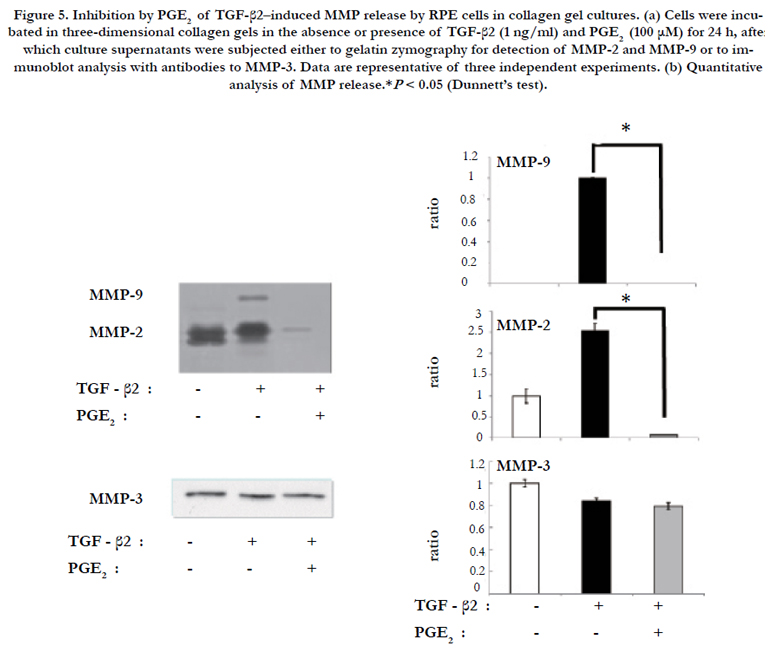
Figure 5. Inhibition by PGE2 of TGF-β2–induced MMP release by RPE cells in collagen gel cultures. (a) Cells were incubated in three-dimensional collagen gels in the absence or presence of TGF-β2 (1 ng/ml) and PGE2 (100 μM) for 24 h, after which culture supernatants were subjected either to gelatin zymography for detection of MMP-2 and MMP-9 or to immunoblot analysis with antibodies to MMP-3. Data are representative of three independent experiments. (b) Quantitative analysis of MMP release.*P < 0.05 (Dunnett’s test).
Discussion
We have here shown that PGE2 inhibited the TGF-β2–induced contraction of a collagen gel mediated by mouse RPE cells. PGE2 also inhibited the phosphorylation of Smad2 and paxillin as well as the expression of α-SMA induced by TGF-β2 in these cells. In addition, the TGF-β2–induced release of MMP-2 and MMP- 9 from RPE cells was attenuated by PGE2. These results thus suggest that PGE2 inhibits TGF-β2–induced collagen contraction mediated by RPE cells by preventing the EMT and focal adhesion formation as well as through down-regulation of MMP expression in these cells. Moreover, these effects of PGE2 may be mediated, at least in part, through inhibition of Smad2 signaling elicited by TGF-β2.
Proliferative tissue that forms on or under the retina during the development of PVR or PDR consists of RPE cells, fibroblasts, glial cells, and macrophages surrounded by extracellular matrix (ECM) [13]. The proliferative fibrocellular membranes formed by these cells can give rise to tractional retinal detachment. The contribution of RPE cells to these disorders is promoted by the EMT, which is characterized in part by the induction of α-SMA and fibronectin and the acquisition of contractile properties [14]. We have now shown that PGE2 inhibited the TGF-β2–induced expression of α-SMA in, as well as collagen gel contraction mediated by, RPE cells. Interactions with the ECM play a key role in the generation of mechanical force during cell contraction [15].
MMPs mediate remodeling of the ECM and thereby contribute to wound healing in the eye as well as to retinal disease, with MMP-2 and MMP-9 having been implicated in the pathogenesis of diabetic retinopathy [16]. MMP-2 has also been shown to play a role in the generation of mechanical stress in RPE cells [17]. We found that the release of MMP-2 and MMP-9 induced by TGF-β2 in RPE cells was inhibited by PGE2. Moreover, we found that a MMPspecific inhibitor suppressed the TGF-β2–induced contraction of RPE cells [18]. These results thus suggest that PGE2 attenuates induction of the EMT as well as MMP expression by TGF-β2 in RPE cells, and that it might therefore also inhibit the formation or contraction of proliferative fibrocellular membranes in PVR or PDR.
Interactions with the ECM increase tractional force in fibroblasts and epithelial cells [19]. Tyrosine phosphorylation of paxillin induced by ECM-cell interaction contributes to the formation of focal adhesions and F-actin reorganization [20]. We have previously shown that tyrosine phosphorylation of paxillin plays role in the adhesion and migration of corneal epithelial cells [21]. Endothelin-1 was also previously shown to increase the contractility of lung fibroblasts by up-regulating the expression of α-SMA, paxillin, ezrin, and moesin [22]. In addition, we previously found that the phosphorylation of paxillin is induced during collagen gel contraction mediated by cultured corneal fibroblasts [23]. Our present results show that PGE2 inhibited paxillin phosphorylation associated with the TGF-β2–induced contraction of RPE cells, suggesting that PGE2 modulates the activation of focal adhesions by TGF-β2 in these cells, and that it might therefore inhibit the contraction of proliferative fibrocellular membranes in individuals with PVR or PDR.
PGE2 is a key regulator of inflammation. However, whereas PGE2-EP4 signaling promotes immune inflammation in various diseases, PGE2 also suppresses the production of inflammatory cytokines by dendritic cells and appears to exert anti-inflammatory effects [24]. The production of PGE2 is up-regulated in certain ocular diseases, suggesting that this agent plays a role in these conditions [25]. We have examine that the EP4 receptor is expressed in the proliferative lesions of PVR or PDR patients (data not shown). These results suggested that this receptor might mediate the inhibitory effect of PGE2 on TGF-β2–induced RPE cell contraction as well as a potential inhibitory effect of PGE2 on the contraction of the proliferative fibrocellular membranes.
In summary, we have shown that PGE2 inhibited TGF-β2– induced collagen contraction mediated by RPE cells. PGE2 also inhibited the expression of α-SMA, the phosphorylation of paxillin, as well as the up-regulation of MMP-2 and MMP-9 in TGF-β2–stimulated RPE cells, with these effects likely resulting at least in part from attenuation of TGF-β2–induced Smad2 signaling. Our results thus suggest that PGE2 warrants further investigation as a potential drug for the treatment of PVR, PDR, and other proliferative retinal diseases.
Acknowledgements
We thank Yukari Mizuno and Shizuka Murata as well as the staff of Yamaguchi University Center for Gene Research for technical assistance.
References
- Hinton DR, He S, Jin ML, Barron E, Ryan SJ (2002) Novel growth factors involved in the pathogenesis of proliferative vitreoretinopathy. Eye 16(4): 422-428.
- Lee H, O'Meara SJ, O'Brien C, Kane R (2007) The role of gremlin, a BMP antagonist, and epithelial-to-mesenchymal transition in proliferative vitreoretinopathy. Invest Ophthalmol Vis Sci 48(9): 4291-4299.
- Breyer RM, Bagdassarian CK, Myers SA, Breyer MD (2001) Prostanoid receptors: subtypes and signaling. Annu Rev Pharmacol Toxicol 41: 661-690.
- Norel X, Walch L, Labat C, Gascard JP, Dulmet E, et al. (1999) Prostanoid receptors involved in the relaxation of human bronchial preparations. Br J Pharmacol 126(4): 867-872.
- Maher SA, Belvisi MG (2010) Prostanoids and the cough reflex. Lung 188(Suppl 1): S9-12.
- Takayama K, Garcia-Cardena G, Sukhova GK, Comander J, Gimbrone MA Jr, et al. (2002) Prostaglandin E2 suppresses chemokine production in human macrophages through the EP4 receptor. J Biol Chem 277(46): 44147-44154.
- Biswas S, Bhattacherjee P, Paterson CA (2004) Prostaglandin E2 receptor subtypes, EP1, EP2, EP3 and EP4 in human and mouse ocular tissues--a comparative immunohistochemical study. Prostaglandins Leukot Essent Fatty Acids 71(5): 277-288
- Parapuram SK, Ganti R, Hunt RC, Hunt DM (2003) Vitreous induces components of the prostaglandin E2 pathway in human retinal pigment epithelial cells. Investigative ophthalmology & visual science 44(4): 1767-1774.
- Yanni SE, Barnett JM, Clark ML, Penn JS (2009) The role of PGE2 receptor EP4 in pathologic ocular angiogenesis. Invest Ophthalmol Vis Sci 50(11): 5479-5486.
- Rocca B, FitzGerald GA (2002) Cyclooxygenases and prostaglandins: shaping up the immune response. Int Immunopharmacol 2(5): 603-630.
- Jo YJ, Sonoda KH, Oshima Y, Takeda A, Kohno R, et al. (2011) Establishment of a new animal model of focal subretinal fibrosis that resembles disciform lesion in advanced age-related macular degeneration. Invest Ophthalmol Vis Sci 52(9): 6089-6095.
- Zhou H, Kimura K, Orita T, Nishida T, Sonoda KH (2012) Inhibition by medroxyprogesterone acetate of interleukin-1beta-induced collagen degradation by corneal fibroblasts. Invest Ophthalmol Vis Sci 53(7): 4213-4219.
- Williams DF, Burke JM (1990) Modulation of growth in retina-derived cells by extracellular matrices. Invest Ophthalmol Vis Sci 31(9): 1717-1723.
- Hinz B, Celetta G, Tomasek JJ, Gabbiani G, Chaponnier C (2001) Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol Biol Cell 12(9): 2730-2741.
- Ramtani S (2004) Mechanical modelling of cell/ECM and cell/cell interactions during the contraction of a fibroblast-populated collagen microsphere: theory and model simulation. J Biomech 37(11): 1709-1718.
- Kowluru RA, Zhong Q, Santos JM (2012) Matrix metalloproteinases in diabetic retinopathy: potential role of MMP-9. Expert Opin Investig Drugs 21(6): 797-805.
- Hou X, Han QH, Hu D, Tian L, Guo CM, et al. (2009) Mechanical force enhances MMP-2 activation via p38 signaling pathway in human retinal pigment epithelial cells. Graefes Arch Clin Exp Ophthalmol 247(11): 1477-1486.
- Kimura K, Orita T, Fujitsu Y, Liu Y, Wakuta M, et al. (2014) Inhibition by female sex hormones of collagen gel contraction mediated by retinal pigment epithelial cells. Invest Ophthalmol Vis Sci 55(4): 2621-2630.
- Saez A, Buguin A, Silberzan P, Ladoux B (2005) Is the mechanical activity of epithelial cells controlled by deformations or forces? Biophys J 89(6): L52-L54.
- Rozengurt E, Rodriguez-Fernandez JL (1997) Tyrosine phosphorylation in the action of neuropeptides and growth factors. Essays Biochem 32: 73-86.
- Teranishi S, Kimura K, Nishida T (2009) Role of formation of an ERK-FAK-paxillin complex in migration of human corneal epithelial cells during wound closure in vitro. Invest Ophthalmol Vis Sci 50(12): 5646-5652.
- Shi-Wen X, Chen Y, Denton CP, Eastwood M, Renzoni EA, et al. (2004) Endothelin-1 promotes myofibroblast induction through the ETA receptor via a rac/phosphoinositide 3-kinase/Akt-dependent pathway and is essential for the enhanced contractile phenotype of fibrotic fibroblasts. Mol Biol Cell 15(6): 2707-2719.
- Liu Y, Yanai R, Lu Y, Kimura K, Nishida T (2006) Promotion by fibronectin of collagen gel contraction mediated by human corneal fibroblasts. Exp Eye Res 83(5): 1196-1204.
- Jing H, Vassiliou E, Ganea D (2003) Prostaglandin E2 inhibits production of the inflammatory chemokines CCL3 and CCL4 in dendritic cells. J Leukoc Biol 74(5): 868-879.
- Bianchi E, Scarinci F, Grande C, Artico M (2012) Immunohistochemical profile of VEGF, TGF-beta and PGE(2) in human pterygium and normal conjunctiva: experimental study and review of the literature. Int J Immunopathol Pharmacol 25(3): 607-615.